RECIPES
Selection of previously unknown structures and description of parameterization used in their solution

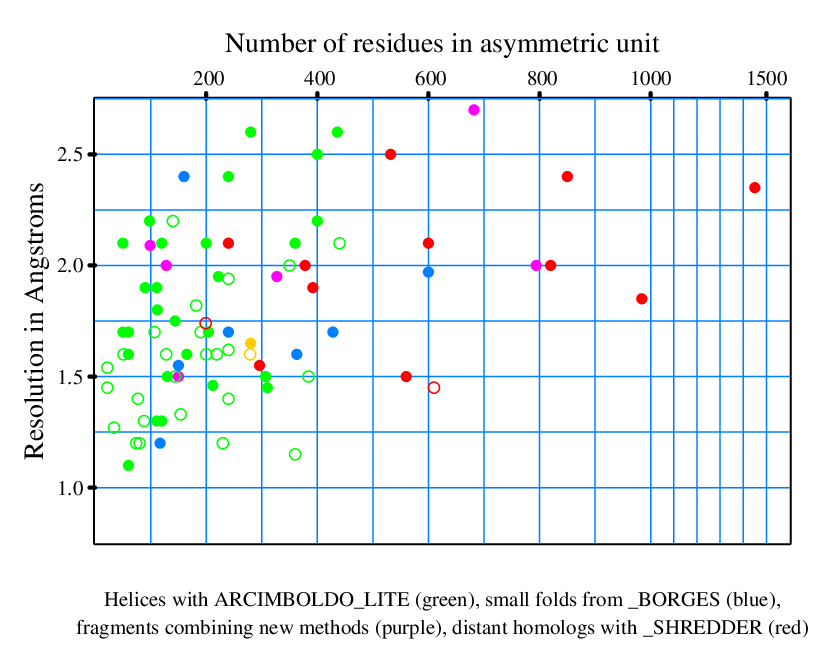
|
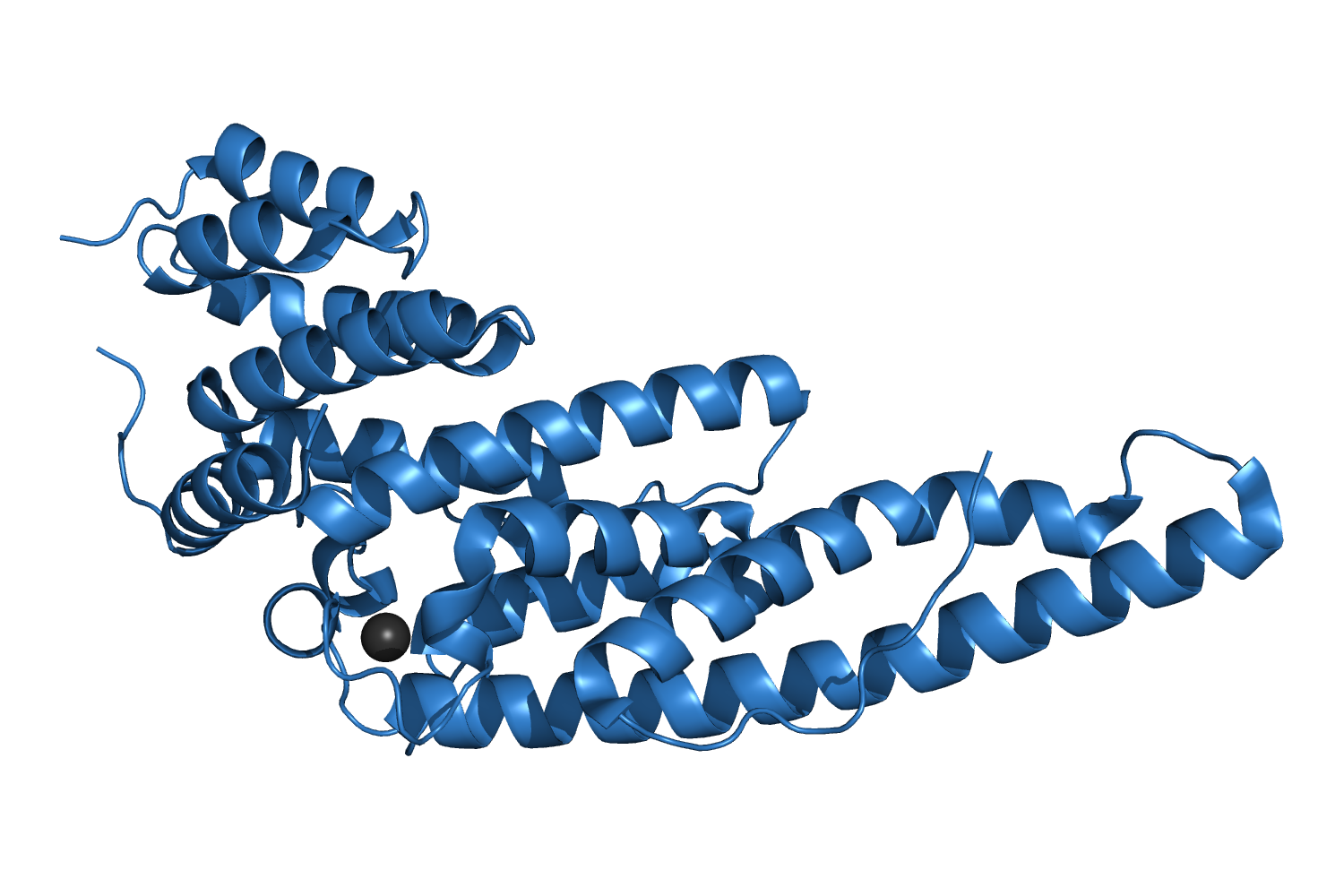
4LUN, A highly conserved region essential for NMD in the Upf2 N-terminal domain Fourati Z, et al., 2014. |
| 310 aa, P21 at 1.45 Å resolution. |
|
| ARCIMBOLDO_LITE, search fragments: 1 ideal helix of 16 alanines originally solved on a grid of 240 cores but solved as well on a machine of 8 cores. |
|
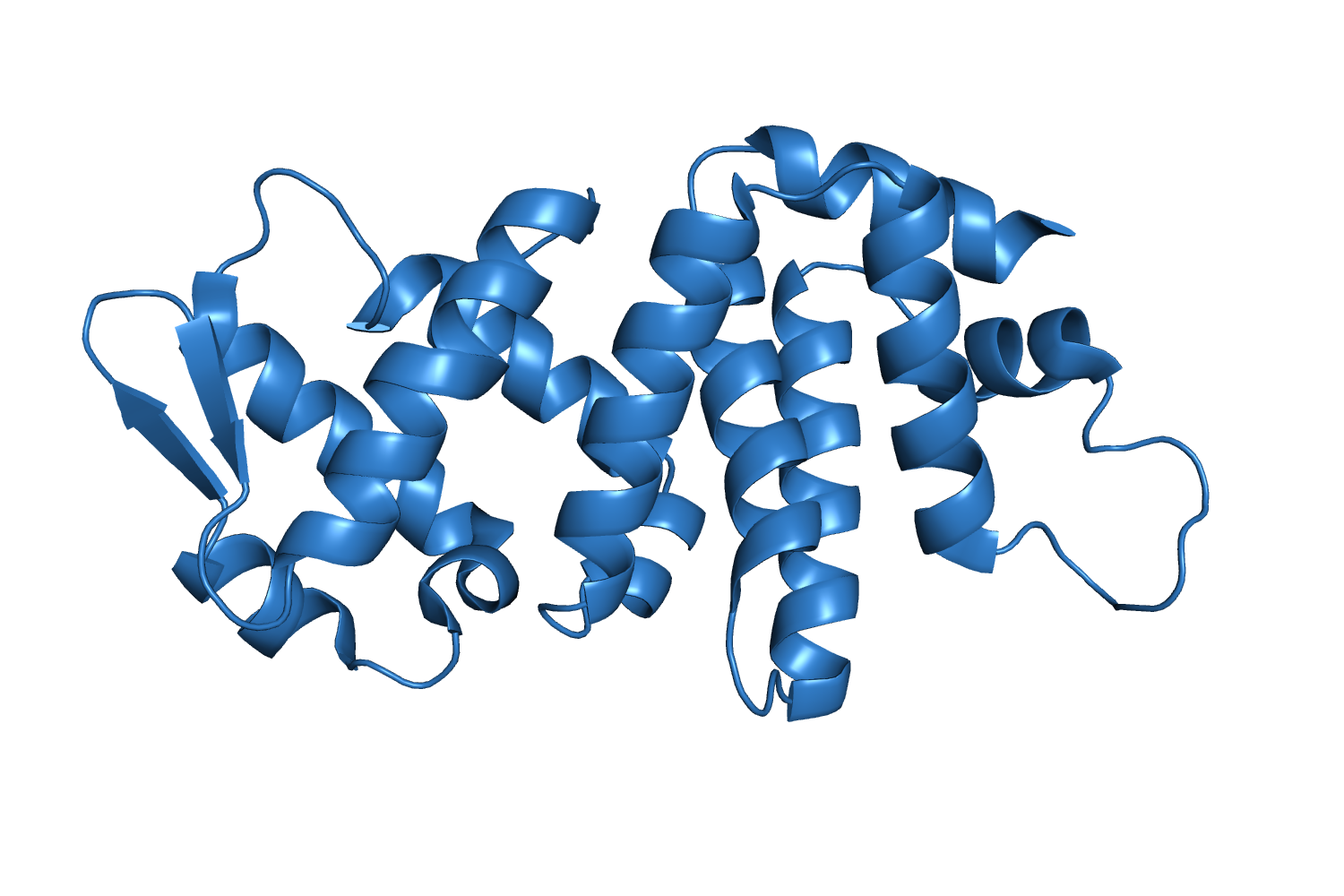
4K82, Unique Crystal Structure of a Novel Surfactant Protein from the Foam Nest of the Frog Leptodactylus vastus Cavalcante Hissa D, et al., 2014. |
| 204 aa, P21 at 1.7 Å resolution. |
|
| ARCIMBOLDO_LITE, search fragments: 2 ideal helices of 14 alanines originally solved on a grid of 240 cores but solved as well on a machine of 8 cores. |
|
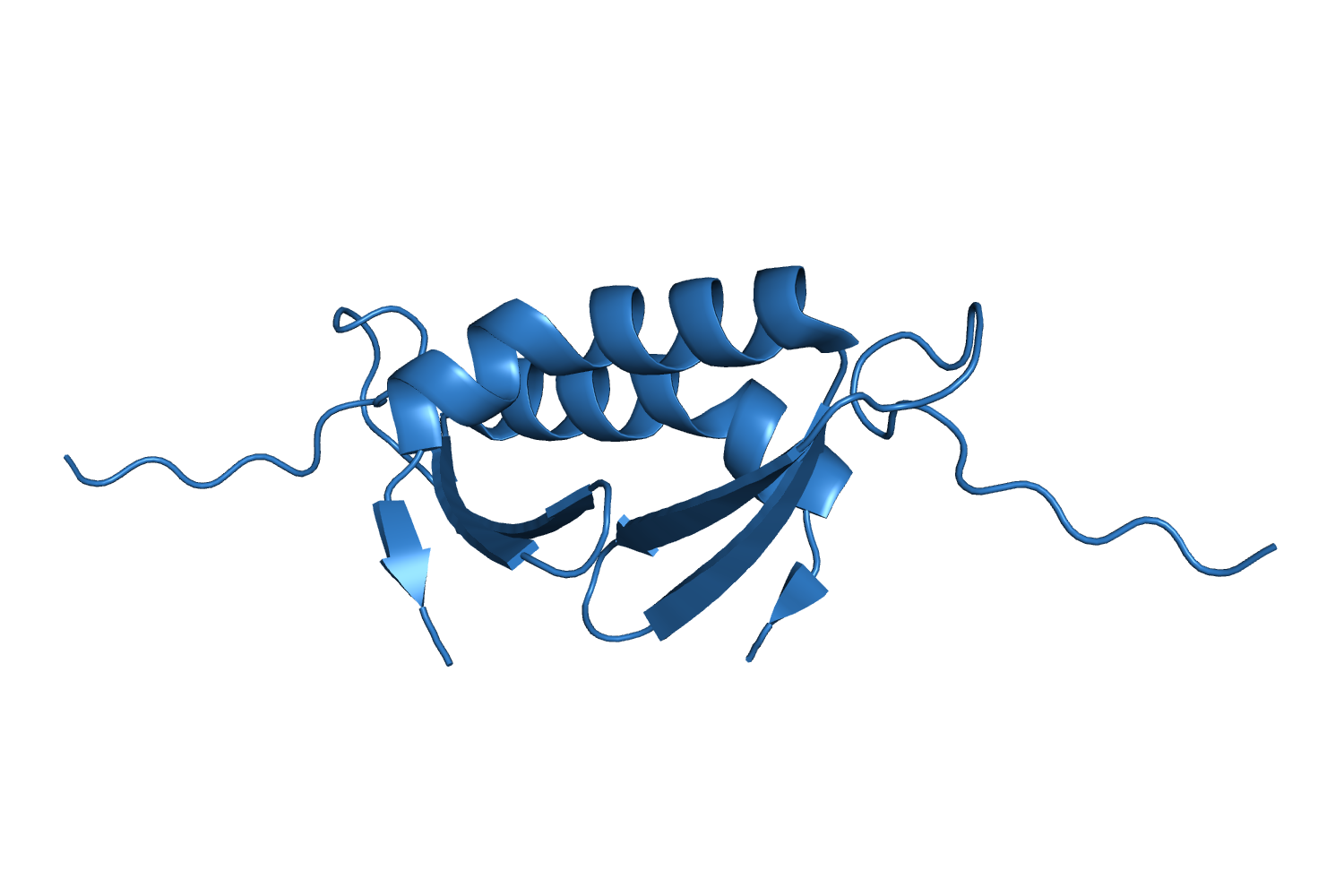
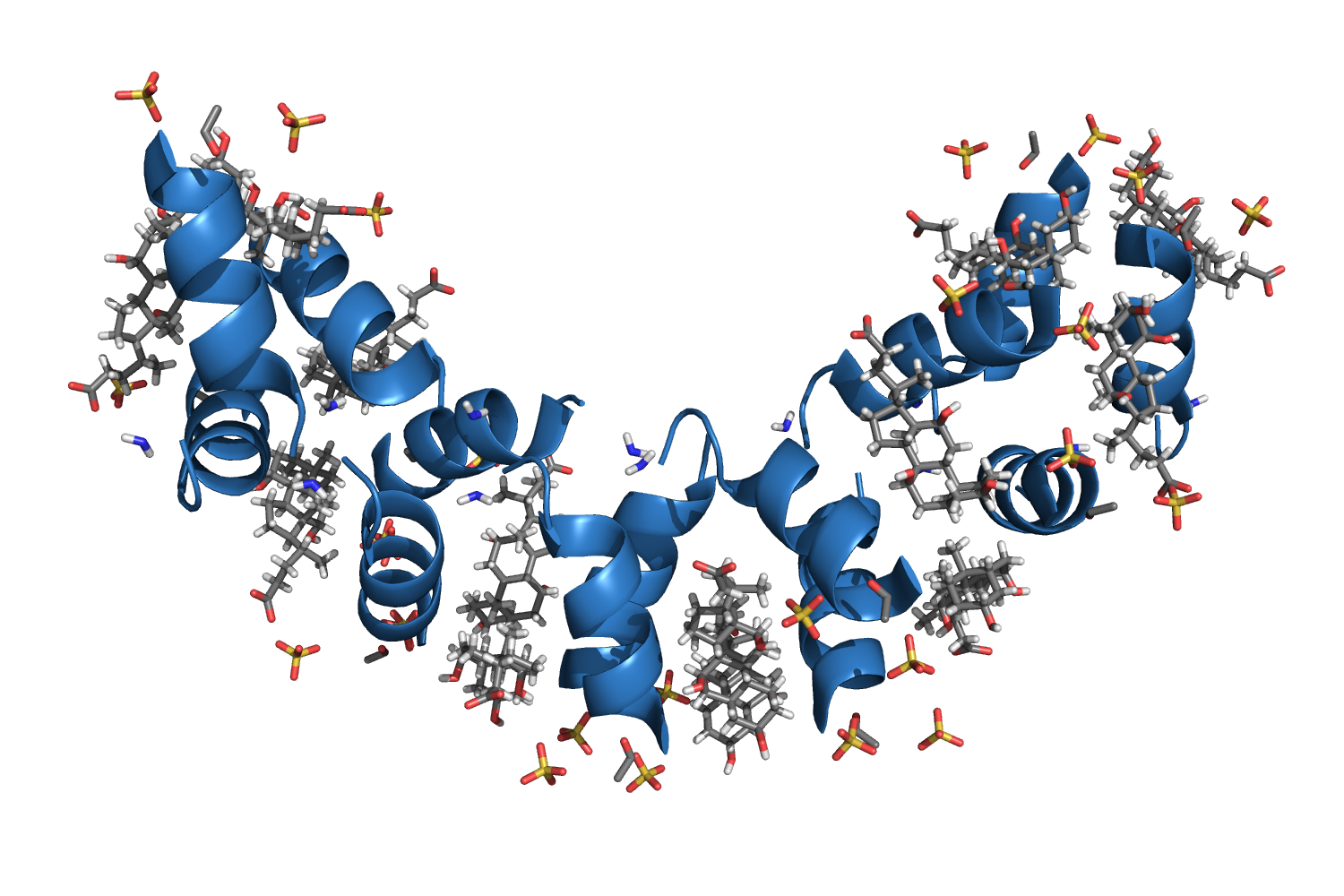
4E1P, The structure of the oligomerization domain of Lsr2 from Mycobacterium tuberculosis reveals a mechanism for chromosome organization and protection Summers EL, et al., 2012. |
| 112 aa, P21 at 1.8 Å resolution. |
|
| ARCIMBOLDO_LITE, search fragments: 2 ideal helices of 12 alanines originally solved on a grid of 240 cores but solved as well on a machine of 8 cores. |
|
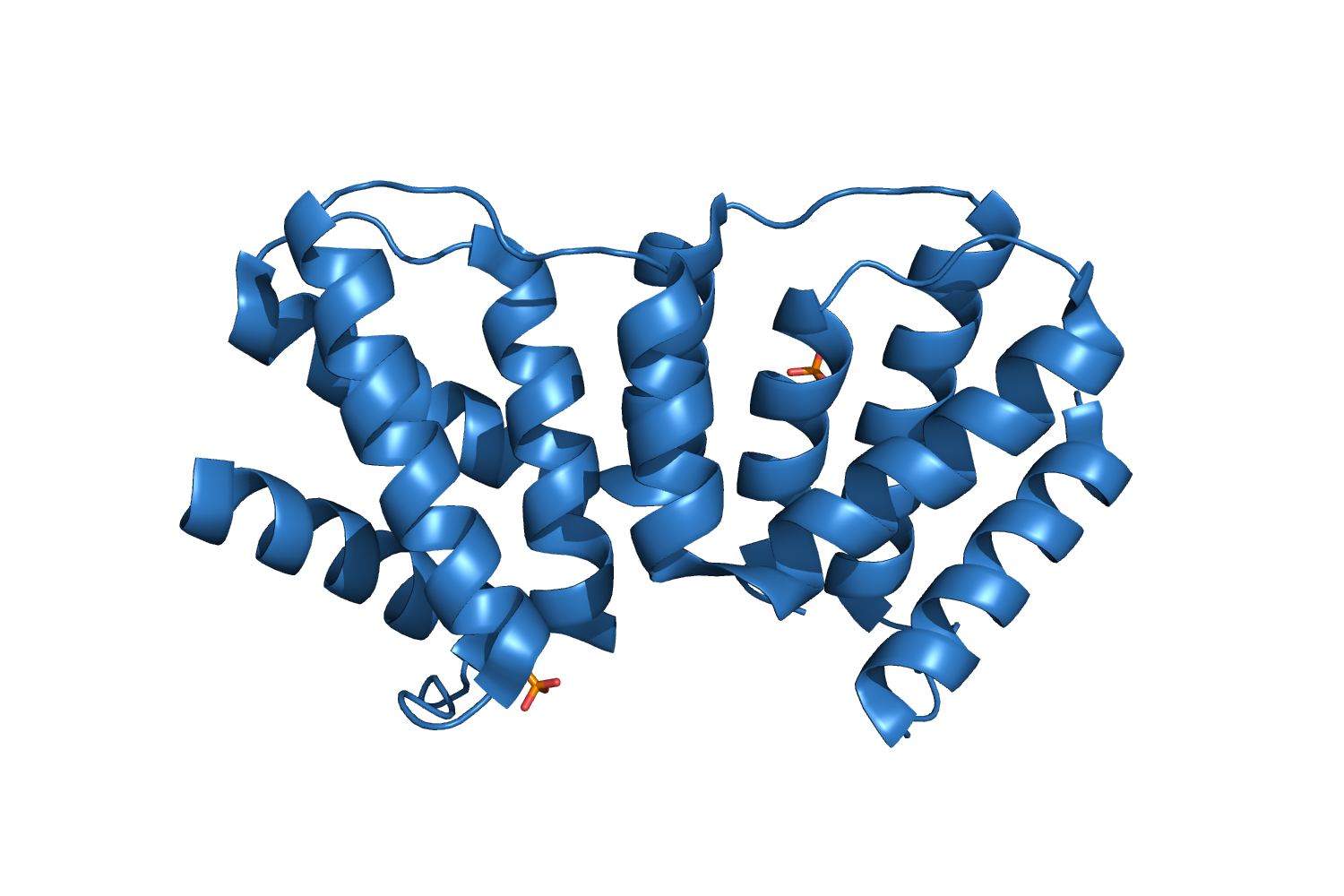
4WG0, Structure of a 13-fold superhelix (almost) determined from first principles Schoch GA, et al., 2015. |
| 182 aa, C2 at 1.82 Å resolution. |
|
| ARCIMBOLDO_LITE, search fragments: 6 ideal helices of 10 alanines solved on a machine of 8 cores. |
|
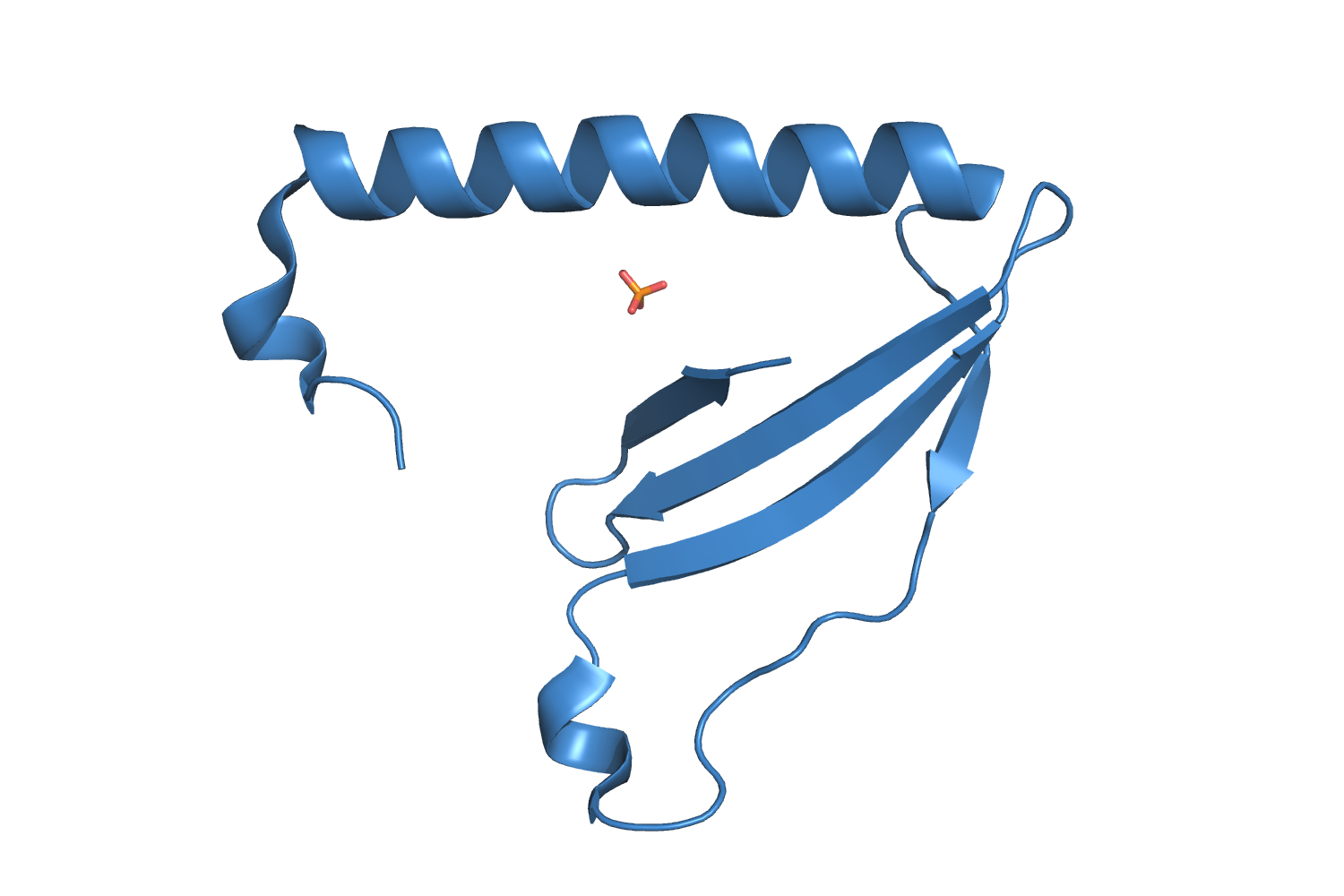
4AEQ, The crystal structure of the dimeric colicin M immunity protein displays a 3D domain swap Usón I, et al., 2012. |
| 90 aa, C2221 at 1.9 Å resolution. |
|
| ARCIMBOLDO_LITE, search fragments: 1 ideal helix of 12 alanines originally solved on a grid of 240 cores but solved as well on a machine of 8 cores.
|
|
3GWH, Crystallographic ab initio protein structure solution below atomic resolution Rodríguez DD, et al., 2009. |
| 222 aa, P21 at 1.95 Å resolution. |
|
| ARCIMBOLDO_LITE, search fragments: 3 ideal helices of 14 alanines originally solved on a grid of 100 cores but solved as well on a machine of 8 cores.
|